 We know a lot about protein synthesis and structure. Proteins are made by the translation machinery (ribosomes + factors) as they copy the information in messenger RNA. When they are first synthesized, proteins can be visualized as random coils or even linear molecules consisting of a long string of amino acids joined end-to-end.
We know a lot about protein synthesis and structure. Proteins are made by the translation machinery (ribosomes + factors) as they copy the information in messenger RNA. When they are first synthesized, proteins can be visualized as random coils or even linear molecules consisting of a long string of amino acids joined end-to-end. Eventually these newly synthesized molecules have to fold into a specific three-dimensional shape that's different for every protein. The diagram on the right illustrates this process for some hypothetical folding pathways.
To a first approximation, the final three-dimensional shape is determined by the amino acid sequence of the protein. The final shape represents the lowest free energy state of the folded protein and this can be represented as a free energy well. Left to their own devices, almost all proteins will eventually reach the bottom of the deepest well that represents the functional state of the protein. (There are exceptions to every rule in biology but this is a very good generality.)
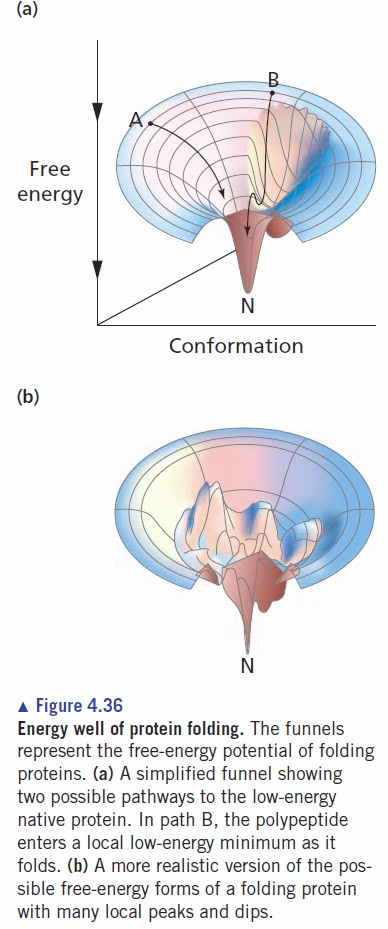 There are many dips and troughs in the free energy landscape and sometimes proteins get trapped in a local minimum as shown by path B in the diagram on the left. If you wait long enough, the incorrectly folded protein will eventually get out of the local dip and fold into the correct shape. (This depends on an energy of activation.)
There are many dips and troughs in the free energy landscape and sometimes proteins get trapped in a local minimum as shown by path B in the diagram on the left. If you wait long enough, the incorrectly folded protein will eventually get out of the local dip and fold into the correct shape. (This depends on an energy of activation.) For the majority of proteins, this spontaneous folding is quite rapid. They reach the proper three-dimensional structure in seconds or minutes. For some proteins it may take much longer, especially if the free energy landscape is rugged and has many deep pits. When a spontaneous biochemical reaction is too slow to be useful it usually means that an enzyme is required to speed up the reaction. Recall that the role of enzymes is to accelerate reactions that occur spontaneoulsy—they do not create new reactions.
The "enzymes" that speed up protein folding are called molecular chaperones and they are among the most highly conserved enzymes in all of biology. As you might expect, these ancient enzymes are present in all species. There are several different kinds of chaperones but one of the most common is called HSP70 (heat shock protein of 70kDa). [Heat Shock and Molecular Chaperones] [The Evolution of the HSP70 Gene Family] [Gene HSPA5 Encodes BiP-a Molecular Chaperone].
HSP70 binds to hydrophobic regions of the folding protein preventing it from aggregating with other partially folded proteins and steering it toward the final three-dimensional structure. This greatly speeds up the folding pathway for those proteins that are otherwise slow to fold. Obviously there has been selection for rapidly folding proteins and/or selection for those that can be effectively assisted by chaperones. The genes for other proteins have not survived so what we see today are proteins that can fold rapidly with, or without, the assistance of chaperones.
Ulrich Hartl has just published a nice review of chaperones in Nature (Hartl et al. 2011). It didn't take long for the IDiots to comment. I spotted a posting on Uncommon Descent:Nature Review Article Yields Unpleasant Data For Darwinism, but that's just a link to another blog posting by a British IDiot named Antony Latham: New research on protein folding demonstrates intelligent design. Here's what Antony Latham has to say about chaperones.
The review in the journal Nature does not discuss the origins of these systems but we need to ask a question: how does all this fit with current evolutionary theory? One might think that such complex systems are confined to mammals or at least the higher orders of animals. This would be a mistake however, because chaperones and chaperonins are in bacteria and archaea also. Indeed it would seem that for any cell to function there needs to be not just proteins but, at the same time, these chaperone systems, which are absolutely essential for proper folding and maintenance of proteins. Without such systems, in place already, the cell will not function.All of the common chaperones fold spontaneously without the assistance of any other chaperones. The reason why they are called "heat shock" proteins is because their synthesis is induced when cells encounter high temperature or other conditions that may cause proteins to unfold or become unstable. These rescue chaperones are made in huge quantities under these conditions to help prevent the destruction of normal cellular proteins. If you understand this then you will understand that the chaperones themselves are capable of rapid spontaneous folding. Even if you didn't know the facts this would seem obvious.
Now, as explained, these chaperone systems are themselves made of proteins which also require the assistance of chaperones to correctly fold and to maintain integrity once folded. Chaperones for chaperones in fact. The very simplest of cells that we know of have these systems in place.
Darwinian evolution requires step by step changes in molecular systems, with one step leading to another in a manner that is statistically reasonable to expect from selection of mutant strains. There is no Darwinian explanation however for the evolution of proteins which already have chaperone systems in place to ensure proper function.
This points very strongly to an intelligent origin of these ‘ingenious’ systems found in all of life.

In the beginning, you didn't need chaperones because every protein folded rapidly on its own. Some of these primitive proteins might have been a bit slow to fold so the evolution of the first chaperones was advantageous because it enhanced the rate of folding for these proteins. The chaperones weren't absolutely necessary for survival but they conferred a selective advantage on those cells that had them.
Once chaperones were present, new proteins could evolve that would otherwise have been too slow to fold in the absence of chaperones. Over time, cells accumulated more and more of these slowly folding proteins so that today no cell can survive without chaperones.
What we can't explain is why the IDiots keep putting their foots in their mouths.
Hartl, F.U., Bracher, A., and Hayer-Hartl, M. (2011) Molecular chaperones in protein folding and proteostasis. Nature 475: 324–332. [Nature]
No comments:
Post a Comment